1. AI applications in ophthalmology
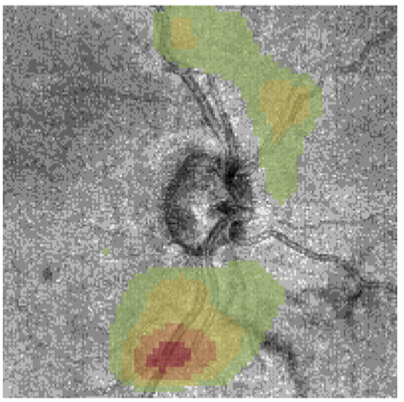 At clinics, monitoring the progression of glaucoma traditionally relies on visual field tests, which is unreliable and time consuming. In collaboration with clinical researchers at NYU, I investigated methods to infer visual field functions directly from scans of the retina, using 3D convolutional neural networks.
At clinics, monitoring the progression of glaucoma traditionally relies on visual field tests, which is unreliable and time consuming. In collaboration with clinical researchers at NYU, I investigated methods to infer visual field functions directly from scans of the retina, using 3D convolutional neural networks.
Publication:
- Yu et al. (in press) Estimating global visual field indices in glaucoma by combining macula and optic disc OCT scans using 3-dimensional convolutional neural networks Ophthalmology Glaucoma [link]
2. Computational neuroscience
The formation of retinotopic maps in the visual cortex
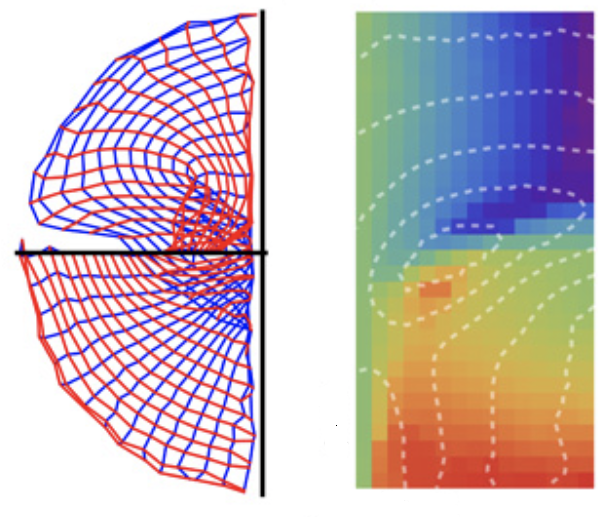 In the visual cortex, modules (called “areas”) are organized as maps. In early visual cortex, adjacent maps tend to be mirror images of each other, but does this organization extend to higher-order areas? In order to understand the general principles of the organization of adjacent maps, I used computational models to study how the development of a retinotopic map could be influenced by its neighbors. The result suggests a novel arrangement can emerge, which can explain unusual features observed in the dorsal visual cortex, which have been puzzling for decades.
In the visual cortex, modules (called “areas”) are organized as maps. In early visual cortex, adjacent maps tend to be mirror images of each other, but does this organization extend to higher-order areas? In order to understand the general principles of the organization of adjacent maps, I used computational models to study how the development of a retinotopic map could be influenced by its neighbors. The result suggests a novel arrangement can emerge, which can explain unusual features observed in the dorsal visual cortex, which have been puzzling for decades.
- Yu et al. (2020) A twisted visual field map in the primate dorsomedial cortex predicted by topographic continuity. Science Advances 6, EAAZ8673. [link]
- A general-audience blog post at the Brain Dialog.
The neural coding of moving patterns
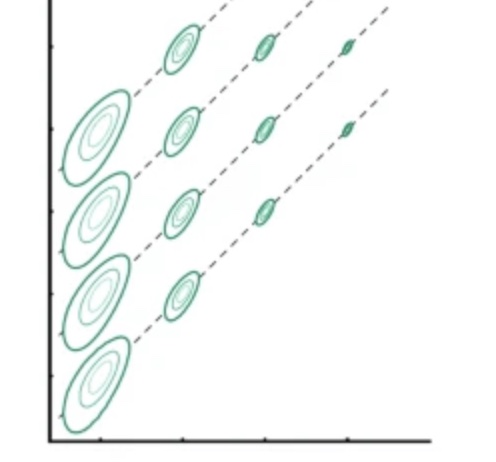 How moving patterns are encoded in the visual cortex is a complex question with many unresolved problems. I examined the ambiguity in the coding of motion speed, and showed that it can be reduced when the object moves in a trajectory.
How moving patterns are encoded in the visual cortex is a complex question with many unresolved problems. I examined the ambiguity in the coding of motion speed, and showed that it can be reduced when the object moves in a trajectory.
Publications:
- Davies, Chaplin, Rosa, Yu (2016) Natural motion trajectory enhances the coding of speed in primate extrastriate cortex. Scientific Reports 6, e19739. [link].
- Yu et al. (2010) Spatial and temporal frequency in striate cortex: functional uniformity and specializations related to receptive field eccentricity. European Journal of Neuroscience 31, 1043-1062. [link].
3. Brain mapping
High-resolution mapping of the controversial primate dorsal visual cortex
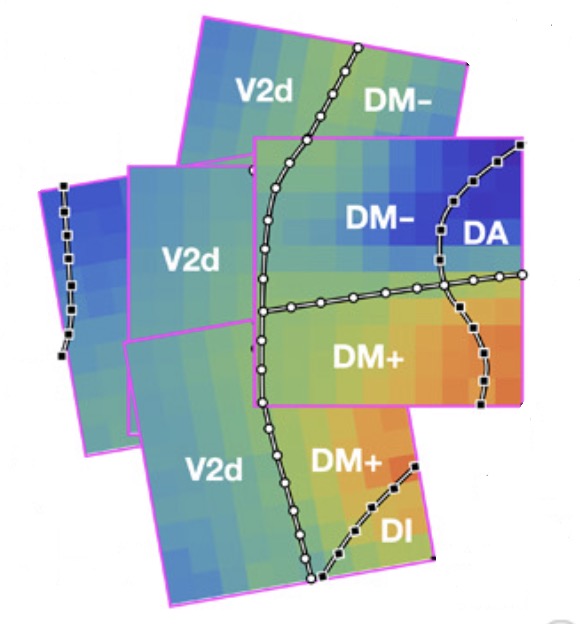 Despite the tremendous efforts invested in creating an atlas for the primate visual cortex, many controversies remain. For example: which area lies rostral to area V2? The conventional answer is area V3, but recent mapping data suggest unexpected complexities (see this review paper and this chapter that I wrote). My colleagues and I mapped the dorsomedial (DM) region of the marmoset visual cortex using high-resolution electrode arrays, and showed that the retinotopy was consistent with the unconventional organization described in Rosa & Schmid (1995), Sereno et al. (2015), and Zhu et al. (2019). I proposed a computational model to explain why this unusual structure might be the natural consequence of computational principles of map organization.
Despite the tremendous efforts invested in creating an atlas for the primate visual cortex, many controversies remain. For example: which area lies rostral to area V2? The conventional answer is area V3, but recent mapping data suggest unexpected complexities (see this review paper and this chapter that I wrote). My colleagues and I mapped the dorsomedial (DM) region of the marmoset visual cortex using high-resolution electrode arrays, and showed that the retinotopy was consistent with the unconventional organization described in Rosa & Schmid (1995), Sereno et al. (2015), and Zhu et al. (2019). I proposed a computational model to explain why this unusual structure might be the natural consequence of computational principles of map organization.
Publication:
- Yu et al. (2020) A twisted visual field map in the primate dorsomedial cortex predicted by topographic continuity. Science Advances 6, EAAZ8673. [link]
A “new” area in the visual cortex: area prostriata
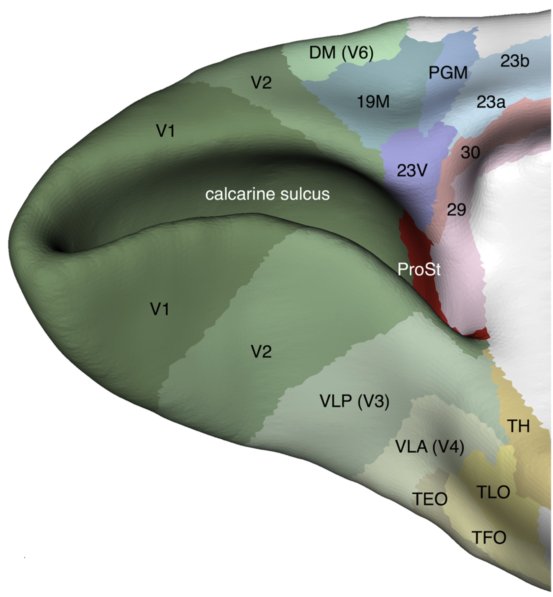 In 2012, my colleagues and I identified a new component of the visual cortex called area prostriata using electrophysiological recordings. This area is weird. It seems to devote most of its resource to the processing of information in the far peripheral visual field (close to the edge of vision), and it seems to be a short-cut for visual information be integrated with non-visual areas in the brain, bypassing most of hierarchy of the visual cortex. A commentary published in Current Biology (Rockland, 2012) stated that this discovery “…raises broad implications for how we view thecortical visual system and,consequently, for how we see”.
In 2012, my colleagues and I identified a new component of the visual cortex called area prostriata using electrophysiological recordings. This area is weird. It seems to devote most of its resource to the processing of information in the far peripheral visual field (close to the edge of vision), and it seems to be a short-cut for visual information be integrated with non-visual areas in the brain, bypassing most of hierarchy of the visual cortex. A commentary published in Current Biology (Rockland, 2012) stated that this discovery “…raises broad implications for how we view thecortical visual system and,consequently, for how we see”.
- My paper: Yu et al. (2012) A specialized area in limbic cortex for fast analysis of peripheral vision. Current Biology 22, 1351-1357. [link]
- The area was confirmed in the human brain using fMRI in 2017. See Mikellidou et al. (2017) Area Prostriata in the Human Brain. Current Biology 27, 3056-3060. [link]
- Commentaries on area prostriata:
- I was interviewed for a Monash university publication about this work.
- Tamietto & Leopold (2018) Visual Cortex: The Eccentric Area Prostriata in the Human Brain. Current Biology 28, pR17–R19. [link]
- Rockland (2012) Visual System: Prostriata — A Visual Area Off the Beaten Path. Current Biology 22, pR571-R573. [link]
- There is (interestingly) a prostriata hashtag on twitter: #prostriata.
4. Neuroplasticity and development
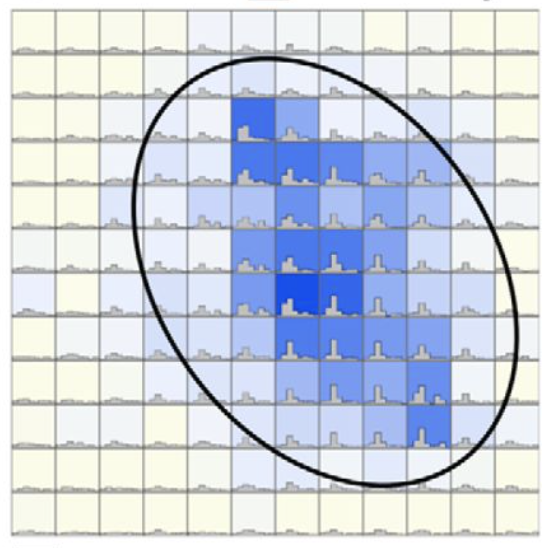 From 2012 to 2017, I conducted a series of studies about how the visual pathways are altered by brain injuries, especially at the early stages of development. Theses studies were designed to elucidate neural pathways for a mysterious phenomenon called blindsight - a condition where patients can react to visual stimuli without being consciously aware of them.
From 2012 to 2017, I conducted a series of studies about how the visual pathways are altered by brain injuries, especially at the early stages of development. Theses studies were designed to elucidate neural pathways for a mysterious phenomenon called blindsight - a condition where patients can react to visual stimuli without being consciously aware of them.
Publications:
- Yu et al. (2018) Robust visual responses and normal retinotopy in primate lateral geniculate nucleus following long-term lesions of striate cortex. Journal of Neuroscience 38, 3955-3970. [link]
- A general-audience summary about the 2018 paper was published in The Brain Dialog.
- Yu et al. (2013) Visually evoked responses in extrastriate area MT after lesions of striate cortex in early life. Journal of Neuroscience 33, 12479-12489. [link]
- Commentary on F1000Prime.
5. Evolution of primate brains
 My collaborator Tristan Chaplin and I used computational methods to compare the brains of primates, in order to answer the following question: do we have big brains because we are smarter? I wanted to publish the result under the title “Larger before Smarter”, because the answer we found was negative. The implications of this finding were discussed in a book chapter I co-authored with Tristan Chaplin.
My collaborator Tristan Chaplin and I used computational methods to compare the brains of primates, in order to answer the following question: do we have big brains because we are smarter? I wanted to publish the result under the title “Larger before Smarter”, because the answer we found was negative. The implications of this finding were discussed in a book chapter I co-authored with Tristan Chaplin.
Publications:
- Chaplin, Yu , Soares, Gattass, Rosa (2013) A conserved pattern of differential expansion of cortical areas in simian primates. Journal of Neuroscience 33, 15120-15125. [link]
- Chaplin, Rosa, Yu (2016) Scaling up the simian primate cortex: a conserved pattern of expansion across brain sizes. In Evolution of Nervous Systems, 2nd edition (Eds. Kaas, JH), Vol. 4, Academic Press. [link]
- The 2016 book chapter was collected in the second edition of Evolutionary Neuroscience, edited by John Kaas. [link]
6. Peripheral vision
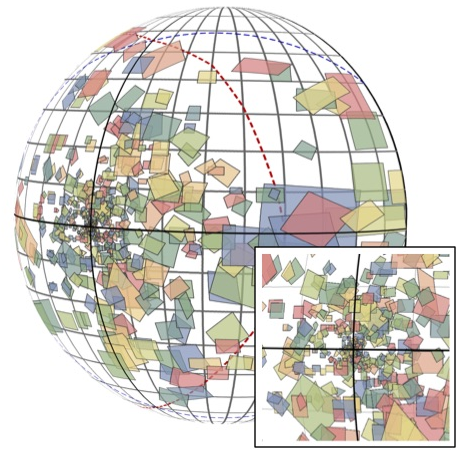 The biggest difference between human vision and computer vision is that human vision is spatially non-uniform: we have a fovea which is specialized for high acuity. But the functional implications of this important difference are not clear. Using electrophysiology, I compared the characteristics of image processing across large expanses of the visual field in the visual cortex.
The biggest difference between human vision and computer vision is that human vision is spatially non-uniform: we have a fovea which is specialized for high acuity. But the functional implications of this important difference are not clear. Using electrophysiology, I compared the characteristics of image processing across large expanses of the visual field in the visual cortex.
Publications:
Yu, Chaplin & Rosa (2015) Representation of central and peripheral vision in the primate cerebral cortex: Insights from studies of the marmoset brain. Neuroscience Research 93, 47-61. [link]
Yu & Rosa (2014) Uniformity and diversity of response properties of neurons in the primary visual cortex: selectivity for orientation, direction of motion and stimulus size from centre to far periphery. Visual Neuroscience 31, 85-98. [link].
Yu et al. (2010) Spatial and temporal frequency in striate cortex: functional uniformity and specializations related to receptive field eccentricity. European Journal of Neuroscience 31, 1043-1062. [link].
7. Computational anatomy and digital atlasing
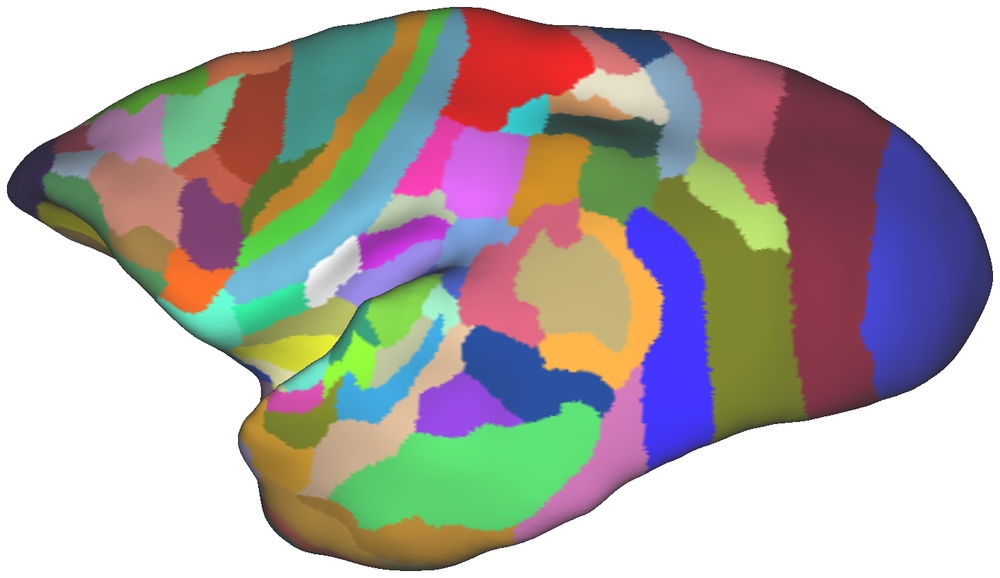 I contributed to figures in the Paxinos marmoset brain atlas, and the early development of the Marmoset Brain Architecture Project.
I contributed to figures in the Paxinos marmoset brain atlas, and the early development of the Marmoset Brain Architecture Project.
Publication:
- Majka, Chaplin, Yu, Tolpygo, Mitra, Wójcik, Rosa (2016) Towards a comprehensive atlas of cortical connections in a primate brain: mapping tracer injection studies of the common marmoset into a reference digital template. Journal of Comparative Neurology 524, 2161-2181. [link]
8. Inventions
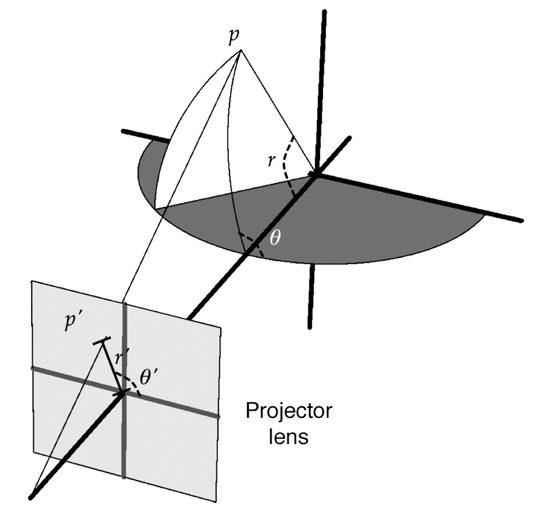 In order to display visual patterns to cover a large proportion of the visual field, I designed and implemented a spherical projection system, and used it extensively in my research. See this method paper.
In order to display visual patterns to cover a large proportion of the visual field, I designed and implemented a spherical projection system, and used it extensively in my research. See this method paper.
At IBM Research, I also filed two USA patents in 2020.